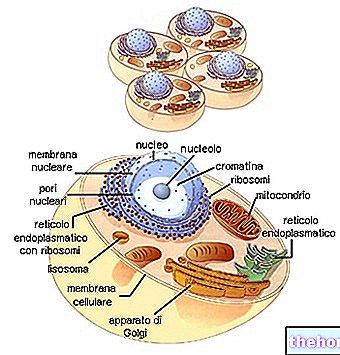
VERBINDUNG (ODER "VERBINDUNG" ODER "VERBINDUNG")
Bisher haben wir die Mendelsche Dihybrid- (oder Polyhybrid-)Kreuzung unter der Annahme behandelt, dass die verschiedenen Allelpaare tatsächlich auf unterschiedlichen homologen Chromosomenpaaren zu finden sind etwa hundert Chromosomen erreichen), während die Zahl der Gene in die Zehntausende gezählt werden kann.
Dass sich die von Mendel für seine Experimente ausgewählten Charaktere alle unabhängig voneinander trennten (ohne damit die Berechnungen der Verteilung der Phänotypen in der F2 der polyhybriden Kreuzung zu verwirren) war ein glücklicher Zufall. Wenn zwei Paare von Allelen an benachbarten Loci gefunden wurden, gilt das Gesetz, dass folgen würde, würde man das Vereinsrecht nennen.
Wenn man weiß, dass sehr viele Charaktere ihren Ort auf einem einzigen Chromosom haben und dass es sich um die Chromosomenpaare handelt, die sich in der Meiose unabhängig voneinander segregieren, kann man gut verstehen, wie häufig es vorkommt, dass zwei Paare von Charakteren vorkommen, wenn sie auf einem Chromosom in der elterlichen Organismus, in gleicher Weise auch in den Gameten und damit in dem Organismus, dem er sein eigenes genetisches Material einbringt, assoziiert bleiben.
So sehen wir, dass „Assoziation“ eine nicht seltene Ausnahme von der Unabhängigkeit darstellt, die in Mendels drittem Gesetz ausgesprochen wird.
AUSTAUSCH ODER "CROSSING-OVER" UND REKOMBINATION
Wenn wir von Meiose sprechen, haben wir darauf hingewiesen, dass es zwei verschiedene Momente der Vermischung des genetischen Materials gibt: Der eine ist der der Chromosomensegregation in den Gameten und der von Mendel beobachtete.
Der andere Moment, der tatsächlich vorausgeht, ist der, in dem die vier Chromatiden jedes Paares homologer Chromosomen gegenseitig identische Merkmale austauschen. Nach diesem Austausch werden zwei Faktoren, die auf demselben Chromosom assoziiert waren, stattdessen in den Gameten unabhängig sein dass ein Austausch stattfindet, ist in erster Näherung proportional zur Länge des Chromosoms, und bei längeren Chromosomen kann es sogar mehr als ein Austausch geben.
Das Phänomen kann zytologisch nachgewiesen werden, indem eine ausreichende Anzahl von Meiose unter dem Mikroskop beobachtet wird.
Die Rematch-Rate ist die Rate, mit der zwei beliebige Charaktere, die in der Elterngeneration assoziiert wurden, in F2 unterschiedlich rekombinieren.
Wenn die beiden Loci absolut zusammenhängend sind, ist die Wahrscheinlichkeit, dass ein Chiasma sie trennt, praktisch null. Die Rekombinationsrate beträgt: n ° Rekombinanten. Wenn sich zwei Loci auf zwei verschiedenen Chromosomen befinden, beträgt die Rekombinationsrate 0,5 (gleiche Wahrscheinlichkeit, dass sich zwei Charaktere, die in der P-Generation verbunden wurden, zufällig in F2 wiederfinden). Die Rekombinationsrate kann daher zwischen 0,0 und 0,5 variieren. Bei kleinen Abständen auf dem Chromosom sind der Abstand und die Rekombinationsrate direkt proportional. Bei längeren Distanzen besteht die Möglichkeit, dass zwei Austausche zwischen zwei Loci stattfinden. Es wird nun klar erscheinen, dass wieder zwei Faktoren, die durch zwei Börsen getrennt sind, miteinander verbunden sind. an dieser Stelle ist klar, dass die Proportionalität zwischen dem Abstand der Loci und der Rekombinationswahrscheinlichkeit verloren geht.
Loci, die auf demselben Chromosom gefunden wurden, bilden "Assoziationsgruppen". Sehr weit entfernte Loci können eine solche Wahrscheinlichkeit der Trennung durch Austausch aufweisen, dass sie sich als unabhängig verhalten, aber jeder von ihnen wird mit einer geringeren Rekombinationsrate mit den dazwischenliegenden Loci assoziiert sein.
Wenn die Rekombinationsraten zwischen vielen Genpaaren innerhalb einer Assoziationsgruppe bekannt sind, kann mit der Erstellung von „genetischen Karten“ begonnen werden. Wenn man bedenkt, dass der Abstand zwischen zwei Genen (a und b) durch die Rekombinationsrate ausgedrückt wird und dass der Abstand von a von einem dritten Gen c entweder die Summe oder die Differenz bezüglich seines Abstands von b sein kann, ist es möglich eine Karte der reziproken Abstände zu rekonstruieren, die die genetische Karte innerhalb dieser Assoziationsgruppe, d. h. dieses Chromosoms, sein wird.
Wir müssen nun allgemein einige Konzepte betrachten, die die phänotypische Manifestation genotypischer Merkmale einschränken.
Zunächst werden wir über die Konzepte der Penetranz und Expressivität sprechen und dann den Phänomenen der Regulation der Genwirkung besondere Aufmerksamkeit widmen.
PENETRANZ
Die Penetranz eines Gens repräsentiert seine Fähigkeit, sich im Phänotyp zu manifestieren. Die Penetranz wird statistisch gemessen, indem die Häufigkeit der Phänotypen, die diesen Charakter aufweisen, von 100 Genotypen, die ihn enthalten, gezählt wird. Ein Merkmal mit einer Penetranz von 0,7 ist ein Merkmal, das in 70 % seiner genotypischen Häufigkeit phänotypisch auftritt.
EXPRESSIVITÄT
Expressivität ist eine quantitative Bewertung des Grades der phänotypischen Manifestation.
REGELUNG DER GENAKTION
Zellen produzieren alle ihre Enzyme und Proteine gleich schnell und gleichzeitig. So können beispielsweise Escherichia coli-Zellen aus dem Lactose-Disaccharid mit Energie und Kohlenstoffatomen versorgt werden, da sie diese dank des Enzyms Beta-Galactosidase in Glucose und Galactose zerlegen können. es gibt ungefähr 3 000 Beta-Galactosidase-Moleküle, was 3 % der Proteine dieser Zelle entspricht; in Abwesenheit von Lactose gibt es nur ein Molekül Beta-Galactosidase pro Bakterienzelle. Galactosidase wird aus neuen mRNA-Molekülen synthetisiert, wenn sie verwendet werden kann. Auch ohne Laktose sind enzymreiche Mutantenstämme von E. coli bekannt: Diese Mutanten sind gegenüber normalen Zellen benachteiligt, da sie zu einem unnötigen Energie- und Materialverbrauch gezwungen sind, das Enzym zu produzieren, das ohne Substrat bleibt. Substanzen, die eine Erhöhung der Enzymmenge bewirken, wie dies bei Laktose der Fall ist, werden als Induktoren bezeichnet, während Enzyme als induzierbar bezeichnet werden. Andere Stoffe induzieren, auch diese auf spezifische Weise, die Produktion bestimmter Enzyme. Auch in E. coli, das beispielsweise alle seine Aminosäuren mit Kohlenstoff und Ammonium (NH3) aufbauen kann, blockiert die Anwesenheit einer bestimmten Aminosäure (z. B. Histidin) im Kulturmedium die Produktion aller damit verbundenen Enzyme die Biosynthese der Aminosäure selbst: Von diesen Enzymen wird gesagt, dass sie unterdrückbar sind.In Bakterienzellen werden die mRNA-Moleküle kurz nach ihrer Bildung abgebaut, und deshalb bedeutet die Kontrolle der Produktion von mRNA gleichzeitig die Kontrolle der enzymatischen Synthese Zeit.
DIE OPERONE
Um zu erklären, wie die Bakterienzelle ihre eigene Produktion von Enzymen kontrollieren kann, formulierten Jacob und Monod die Hypothese des Operons; das Operon wird von mehreren Genen gebildet, die funktionell verwandt und ohne Diskontinuität entlang eines DNA-Abschnitts ausgerichtet sind.Das Operon besteht aus drei verschiedenen Arten von Genen: dem Promotor, an dem die Bildung der mRNA beginnt; der Operator, bei dem die Kontrolle ausgeübt wird; ein oder mehrere Strukturgene, die für Enzyme oder andere Proteine kodieren. Im Beta-Galactosidase-System umfasst das Operon zusätzlich zu dem für Beta-Galactosidase auch zwei weitere Gene, die strukturell kodieren für andere Enzyme, die am Laktosestoffwechsel beteiligt sind.Diese Gene liegen nebeneinander und werden nacheinander entlang derselben DNA-Helix in ein einziges mRNA-Molekül transkribiert.Die so hergestellten mRNA-Moleküle sind für sehr kurze Zeit aktiv und werden danach von bestimmten Enzymen zerstört.
Die Aktivität des Operons wird wiederum von einem anderen Gen gesteuert, dem Regulator, der auch vom Operon entfernt sein kann: Dieser Regulator kodiert für ein Protein, einen sogenannten Repressor, der an die DNA des Operatorgens zu binden scheint zwischen dem Promotor und den Strukturgenen blockiert tatsächlich die Produktion von mRNA.
Der Repressor wiederum wird kontrolliert, und die Kontrolle erfolgt mittels einer "Signal"-Substanz. Bei induzierbaren Enzymen ist diese Substanz der „Induktor. Der Induktor“ bindet an das Repressormolekül und verändert seine Form, sodass es sich nicht mehr an die DNA anpassen kann: in diesem Fall, da zwischen Promotor und Strukturgenen kein Repressor vorhanden ist , kann der Repressor die mRNA-Moleküle und aus diesen die Proteinmoleküle bilden. Wenn der Vorrat an Induktor wieder erschöpft ist, erhält der Regulator die Kontrolle zurück, was die Produktion neuer mRNA, also neuer Proteine, stoppt.Im Beta-Galactosidase-System ist der Induktor Lactose oder eine dieser sehr ähnliche Substanz wird sich dem Repressor anschließen und ihn inaktivieren, um die Biosynthese von Enzymen zu ermöglichen. Bei reprimierbaren Enzymen wirkt die als „Signal“ fungierende Substanz als Co-Pressor: Der Repressor ist nur in Kombination mit dem Co-pressor aktiv. Im Histidin-System, an dem ein Dutzend verschiedener Enzyme beteiligt sind, ist diese Aminosäure in Kombination mit ihrer tRNA der Corepressor, Histidin.
ALLESTERISCHE INTERAKTIONEN
Allosterische Interaktionen, bei denen ein Enzym durch Veränderung seiner Form inaktiviert wird, bieten eine andere Möglichkeit, die Stoffwechselaktivität einer Zelle zu regulieren. Allosterische Wechselwirkungen ermöglichen eine genauere Kontrolle als das Induktor-Repressor-System des Operons, erzielen jedoch nicht das nützliche Ergebnis, die Biosynthese einer bestimmten Substanz aus der ersten Stufe - der Produktion einer mRNA - auszuschließen.
STEUERSYSTEME IM EUKARIUS
Es gibt einige Tatsachen, die zu der Annahme führen, dass ein dem Operon ähnliches Regulationssystem funktioniert und bei Pflanzen und Tieren überragend ist.Die Chromosomen dieser Organismen unterscheiden sich grundlegend von denen von E. coli und anderen Prokaryoten Zellen sind sehr unterschiedlich: Der Mechanismus der Mitose ist so, dass jede Zelle einer bestimmten Pflanze oder eines bestimmten Tieres alle Informationen besitzt
Genetik, die in der befruchteten Eizelle vorhanden ist. Daher bleiben die meisten Gene in jeder spezialisierten Zelle während der gesamten Lebensdauer der Zelle ineffizient. Die DNA in diesen Zellen ist immer mit Proteinen verbunden. Daher ist es möglich, dass die Genrepression in Eukaryoten genau diese Verbindung erfordert zwischen DNA und Proteinen.