Die Mitose wird üblicherweise in vier Phasen unterteilt, die als Prophase, Metaphase, Anaphase und Telophase bezeichnet werden. Ihnen folgt die Teilung in zwei Tochterzellen, die als Zytodierese bezeichnet wird.
Prophase
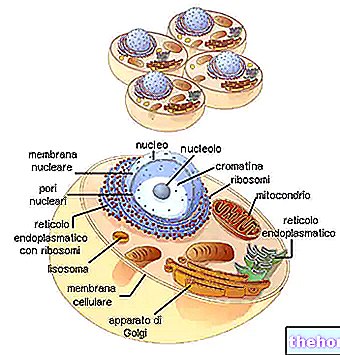
Im Kern sieht man nach und nach die Abgrenzung von färbbaren Filamenten, die noch verlängert und zu einer Kugel gewickelt sind. Die allmähliche Spiralisierung der an die Kernproteine gebundenen DNA-Stränge macht so die Chromosomen allmählich identifizierbar. Währenddessen verschwindet der Nukleolus, während sich das Zentriol verdoppelt. Die beiden Zentriolen wandern zu entgegengesetzten Kernpolen, während die Auflösung der Kernmembran beginnt. Im Moment des Übergangs von der Prophase in die Metaphase (da einige separat als Prometaphase identifiziert werden) sind die Chromosomen verkürzt und deutlich sichtbar, nicht mehr in der Kernmembran isoliert; die Zentriolen befinden sich an entgegengesetzten Polen, mit einer Spindel von Mikrotubuli, die sie mit Meridianformen verbindet: Der Nukleolus hat sich aufgelöst. Die Spindel, die die Zentriolen verbindet, genannt achromatische Spindel, weil sie nicht gefärbt werden kann (im Gegensatz zu den Chromosomen), umfasst sowohl kontinuierliche Fasern (Spindelfasern) als auch Fasern, die in ihrer Mitte mit den Zentromeren der Chromosomen verbunden sind (chromosomale Fasern).

Metaphase
In der Metaphase wird die Spindel deutlich unterschieden, wobei alle Chromosomen auf der Äquatorebene, der sogenannten Äquatorialplatte, angeordnet sind. In diesem Moment sind die Chromosomen maximal verkürzt. Dies ist der Moment, in dem die Zelle darauf fixiert ist, die Chromosomen zu zählen und zu identifizieren. Jedes Chromosom scheint eindeutig aus zwei identischen Filamenten (Chromatiden) zusammengesetzt zu sein, die in einem Punkt namens Zentromer ( die beiden Chromatiden sind das Ergebnis einer Verdoppelung.) Das Zentromer ist der einzige Kontaktpunkt, wie eine Art Adhäsion zwischen zwei Blutplättchen. Alle Zentromere sind am Mittelpunkt der chromosomalen Fasern der achromatischen Spindel befestigt (daher befinden sich die Chromosomen in äquatorialer Position).
Anaphase
Am Ende der Metaphase wird festgestellt, dass sich jedes Zentromer verdoppelt, wobei jede Hälfte entlang der Spindel in Richtung ihres jeweiligen Pols wandert. An dieser Stelle trennen sich die Chromatiden, von ihren jeweiligen Zentromeren gezogen, deutlich in zwei Gruppen: Jedes so getrennte Chromatid ist sozusagen erwachsen geworden: von nun an ist es ein Chromosom, das für die jeweilige Tochterzelle bestimmt ist.
Telophase
Die in zwei identische Cluster getrennten Chromosomen kehren zur Entspiralisierung zurück und bilden den Kern von zwei neuen Zellen; die achromatische Schmelze löst sich auf.
Zytodierese
Auch das Zytoplasma teilt sich durch allmähliche Drosselung, wobei den beiden Tochterzellen die jeweiligen Anteile von Volumen und Zellorganellen zugeschrieben werden. Insbesondere muss jede Tochterzelle mindestens ein Mitochondrium erhalten, da sie ein Zentriol erhalten hat (dies sind, wie erwähnt, Strukturen mit eigener genetischer Kontinuität).
Es sollte beachtet werden, dass im Pflanzenreich, obwohl die allgemeinen Merkmale der Mitose respektiert werden, einige Unterschiede bestehen. Zunächst fehlen die Zentriolen: An den Polen der Spindel befinden sich optisch leere Räume, sogenannte Zentrosomen, von denen die Mikrotubuli ausgehen. Weiterhin erfolgt zum Zeitpunkt der Zytodierese, bei der auch die Zuordnung eines Plastiden (aufgrund seiner genetischen Kontinuität) zu jeder Tochterzelle sichergestellt werden muss, die Trennung der Tochterzellen nicht durch Strangulation, sondern durch Bildung eines Septums, zunächst nur von Plasmalemma, dann mit anschließender Zwischenschaltung der Zellwand.
Die genetische Grundlage stellt der "Wechsel zwischen einer Verdoppelung des Erbguts (Verdoppelung der DNA, d der beiden Chromatiden in die entgegengesetzte Richtung, um die beiden neuen gleichen Kerne zu bilden).
Da, wie wir sehen werden, die Chromosomen in Paaren von Homologen vorliegen (die jeweils von den Gameten stammen), sehen wir, dass die Anzahl der Chromosomenstränge unmittelbar nach der Teilung ein Paar für jeden Chromosomentyp beträgt. Nennt man n die Anzahl der verschiedenen Chromosomentypen, die für die einzelne Spezies charakteristisch sind, beträgt der normale Chromosomensatz nach der Mitose n Paare homologer Chromosomen (2n Chromosomen = diploide Zelle).
Nach der S-Phase hat sich jedoch jedes Chromosom verdoppelt. Um jeder Tochterzelle 2n Chromosomen geben zu können, sind 4n Chromatiden erforderlich. Auf diese Weise sehen wir, dass Reduplikation und Mitose zwischen 4n und 2n DNA-Strängen abwechseln.