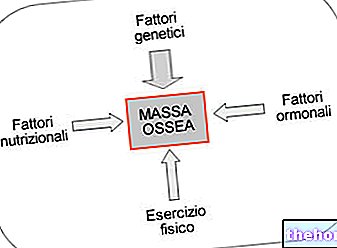
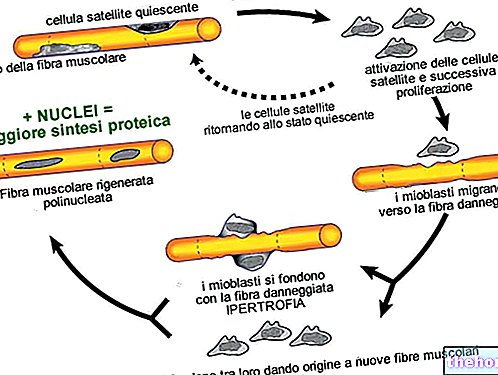
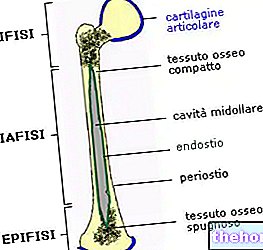
" erster Teil
Das Enzym, das die vierte Stufe des Krebszyklus katalysiert, ist das α-Keto-Glutarat-Dehydrogenase; Dieses Enzym ist ein Enzymkomplex, der der Pyruvatdehydrogenase sehr ähnlich ist. Beide bestehen aus 48-60 Proteinen, in denen drei verschiedene enzymatische Aktivitäten erkannt werden und haben auch die gleichen enzymatischen Cofaktoren; sind sehr ähnliche Enzyme, weil sie auf ähnliche Substrate wirken: sowohl Pyruvat als auch l "α-Ketoglutarat, sind α- Ketosäuren. Der Wirkmechanismus der beiden enzymatischen Komplexe ist der gleiche.
Der Angriff von Thiaminpyrophosphat auf das Carbonyl (C = O) des "α-Ketoglutarat, führt zu seiner Decarboxylierung und das Carboxyhydroxypropyl-Derivat wird gebildet. Mit der anschließenden Übertragung auf das Lipoamid findet ein interner Redoxprozess statt, aus dem das Lipoamid-Carboxy-Derivat bzw. Succinyl-Lipoamid gewonnen wird.
Das Succinyllipoamid reagiert dann mit Coenzym A zu Succinylcoenzym A (das sich im Krebszyklus fortsetzt) und dem reduzierten Lipoamid, das durch das FAD reoxidiert wird: das gebildete FADH2 wird durch NAD + reoxidiert und NADH wird erhalten . In dieser Stufe fand daher die zweite Abspaltung eines Kohlenstoffs aus dem kohlenstoffhaltigen Gerüst in Form von Kohlendioxid statt.
Eine an Coenzym A gebundene Acylgruppe liegt in aktivierter Form vor, d. h. sie hat einen hohen Energiegehalt: Es ist daher möglich, die Energie von Succinyl-Coenzym A zu nutzen.

In der fünften Stufe des Krebszyklus wird Succinyl-Coenzym A der Wirkung von Succinylthiokinase; Über seine Wirkungsweise wurden zwei Hypothesen aufgestellt: Wir werden nur eine der beiden beschreiben, da sie die am meisten anerkannte ist. Nach dieser Hypothese wird Succinyl-Coenzym A vom Stickstoff eines Histidins (Hys) des Enzyms angegriffen: Coenzym A wird freigesetzt und als Zwischenprodukt entsteht ein von Histidin abgeleitetes Addukt, das Succinyl-Enzym (oder Succinyl-Hys ); auf dieses Zwischenprodukt wirkt ein Orthophosphat, das zur Freisetzung von Succinat und zur Bildung des Phosphoenzyms führt.Das Phosphoenzym, das von Guanosindiphosphat (GDP) angegriffen wird, produziert Guasnosintriphosphat (GTP) und das Enzym wird freigesetzt. Aus energetischer Sicht GTP = ATP: Die energieliefernde Bindung ist bei beiden Spezies gleich (es ist die Anhydridbindung zwischen Phosphoryl Β und Phosphoryl γ). In einigen Fällen wird GTP als Material mit hohem Energiegehalt verwendet, aber normalerweise wird GTP durch die Wirkung des Enzyms in ATP umgewandelt Nukleosiddiphosphokinase (NDPK); ist ein Enzym, das in Zellen vorkommt und folgende Reaktion katalysiert:
N1TP + N2DP → N1DP + N2TP
Generisches NiTP ® Nucleosid Triphosphate
Generisches NiDP ® Nukleosiddiphosphat
Es ist eine reversible Reaktion; In unserem Fall passiert es:
GTP + ADP → BIP + ATP
daher kann es auch bei kleinen Konzentrationsschwankungen der Reagenzien nach rechts oder links gehen.
Wenn der Krebszyklus so schnell abläuft, dass eine über dem Energiebedarf liegende ATP-Produktion entsteht, ist ADP knapp verfügbar, während viel ATP vorhanden ist: Die von der Nukleosid-Diphospho-Kinase katalysierte Reaktion ist dann nach links gerichtet (GTP akkumuliert, wenn die Nukleosiddiphosphokinase nicht genügend Substrat, d. h. ADP, hat). GTP ist somit ein Signal der Energieverfügbarkeit und verlangsamt somit den Krebszyklus.
Die sechste Stufe des Krebszyklus führt zur Bildung des Fumarats durch die Wirkung von Succinat-Dehydrogenase; dieses Enzym gibt eine stereospezifische Reaktion, da immer das ungesättigte (es ist ein Alken) trans gebildet wird, dh das Fumarat (während das cis-Isomer das Maleat ist). Succinat-Dehydrogenase befindet sich auf der inneren Mitochondrienmembran, während alle anderen Enzyme des Krebs-Zyklus über das Mitochondrium verstreut sind.
Succinatdehydrogenase hat FAD als Cofaktor; es wird durch Oxalacetat (Feedback-Hemmung) gehemmt, während es Succinat und Fumarat als positiven Modulator (Aktivator) hat. seinen Aktivator. Versuchen wir zu verstehen, warum, indem wir in die letzte Phase des Krebszyklus springen Der Krebs-Zyklus benötigt Energie, daher besteht die einzige Möglichkeit, Oxalacetat vom Patienten zu gewinnen, darin, dass die Konzentration des Patienten sehr hoch ist: Malat ist einer der Metaboliten mit der höchsten Konzentration in den Zellen.Die Reaktion, die Malat in Oxalacetat umwandelt, wird auch durch die Tatsache, dass die Konzentration von Oxalacetat durch die Wirkung der Citratsynthase niedrig gehalten wird. Die durch die Succinat-Dehydrogenase katalysierte Reaktion ist also eine Selbstfütterungsreaktion und nur so kann die Umwandlung von Malat in Oxalacetat erfolgen.
Die Konzentration von mitochondrialem Malat muss mit der Konzentration des zytoplasmatischen Malats kompatibel sein: Nur wenn die Konzentration von mitochondrialem Malat so hoch ist, dass die Umwandlung des Malats in Oxalacetat (im Krebs-Zyklus) gewährleistet ist, kann das Malat auch in andere Wege (die zytoplasmatisch sind): Im Zytoplasma kann das Malat in Oxalacetat umgewandelt werden, aus dem durch die Wirkung von GOT (es ist eine Transaminase) oder Glukose durch Gluconeogenese Aspartat gewonnen werden kann.
Wir kehren zur siebten Stufe des Krebszyklus zurück, die durch das Enzym katalysiert wird fumarasi: Wasser wird auf stereospezifische Weise hinzugefügt, um L-Malat herzustellen.
In der letzten Phase des Krebs-Zyklus, von der wir bereits gesprochen haben, ist die Wirkung der Malatdehydrogenase. Dieses Enzym verwendet ein NAD + -Molekül für seine katalytische Wirkung.
Damit haben wir die Beschreibung der verschiedenen Stadien des Krebszyklus abgeschlossen.
Der Krebs-Zyklus ist vollständig reversibel.
Um die Geschwindigkeit des Krebszyklus zu erhöhen, kann die Konzentration der in diesem Zyklus vorhandenen Metaboliten erhöht werden; Eine der Strategien zur Beschleunigung des Krebszyklus besteht darin, einen Teil des Pyruvats, das in die Mitochondrien gelangt, in Oxalacetat umzuwandeln (durch die Wirkung der Pyruvat-Carboxylase) und nicht alles in Acetyl-Coenzym A umzuwandeln: Dadurch erhöht sich die Konzentration von Oxalacetat, das ist ein Metabolit des Krebs-Zyklus und erhöht daher die Geschwindigkeit des gesamten Zyklus.
Im Krebs-Zyklus werden drei NAD+ in drei NADH und ein FAD in FADH2 umgewandelt und außerdem ein GTP gewonnen: Durch die Kanalisierung der Reduktionskraft aus dem Krebs-Zyklus wird weiteres ATP produziert; In der Atmungskette wird die Reduktionskraft von NADH und FADH2 auf Sauerstoff übertragen: Diese Übertragung erfolgt durch eine Reihe von Enzymen, die sich auf der Mitochondrienmembran befinden und in ihrer Wirkung zur Produktion von ATP führen.
Die Prozesse der Atmungskette sind exergonische Prozesse und die freigesetzte Energie wird zur Produktion von ATP verwendet; der Zweck der Zelle besteht darin, die exergonischen Prozesse zu nutzen, um die Synthese von ATP zu ermöglichen. Für jedes NADH-Molekül, das in die Atmungskette eintritt, werden 2,5 Moleküle ATP erhalten und für jedes FADH2 werden 1,5 Moleküle ATP erhalten; Diese Diversität ist darauf zurückzuführen, dass FADH2 auf einem niedrigeren Niveau in die Atmungskette eindringt als NADH.
Mit der Reduktionskraft des aeroben Stoffwechsels werden 30-32 ATP (219-233 kcal / mol) mit einer Effizienz von etwa 33% erhalten (die Effizienz des anaeroben Stoffwechsels beträgt etwa 2%).